
Professor Daniel Szymanski – Math Plus Biology: Building A Knowledge Base To Engineer Plant Traits
Novel research seeks to unravel one of the most complex mysteries of plant biology and pave the way toward better, denser crops, under the careful guidance of Professor Daniel Szymanski at Purdue University.

Looking into Leaf Growth
Trapped within a thick canopy formed by the leaves of their neighbours, plants must compete for limited sunlight that is used to drive photosynthesis. Plants are able to tune their overall architecture to suit environmental conditions, and produce leaves with optimal sizes, shapes and angles. Leaf development is highly complex, as the leaf bud is converted from a cylindrical peg, into a thin, yet mechanically stable, bladelike structure.
Leaf shape is of utmost importance in agriculture. For example, more erect leaf angles have enabled greater planting densities and higher yields per acre for maize. This trait has been achieved through traditional plant breeding and selection. In the future, there will be opportunities to combine breeding and selection with genetic engineering to generate new types of crops with optimised whole plant architectures. Presently this is not possible however, as we lack sufficient knowledge about how genes, cells, and tissues interact across wide spatial scales to control the overall morphology of the leaf. Therefore, there is a strong need to develop new experimental methods to analyse gene function at the levels of cell, tissue sector, and whole organ.
Professor Szymanski’s research team and collaborators investigate how protein-based molecular machines give rise to cells of particular shapes and sizes, and explore how this can affect macroscopic traits. Specifically, Professor Szymanski’s area of expertise is the genetics and cell biology of plant growth. His research uses the model plant Arabidopsis for molecular genetics, cell biology, and computational biology studies. In addition, some of his newer projects involve legumes, cotton and maize.
Discovering the Cellular Basis of Leaf Shape
In the early stages of leaf development, cylindrical leaf primordia are converted into flat surfaces with polarised shapes. The size and shape of the leaf is determined by the number of cells, and the final sizes and shapes of the cells. Plant cells, unlike animal cells, can increase in volume by a factor of 100 or more after they stop dividing.
Therefore, the magnitude and direction of cell expansion can exert a strong influence on the size and shape of the whole leaf. In plants, the outer layer of cells – termed the epidermis – exerts a strong influence over the growth properties of the organ. In many important crop species, the leaf epidermis is comprised of pavement cells that can adopt wildly varying shapes: some of them are polygonal, while others are highly lobed with finger-like protrusions to generate tissue comprised of interdigitated cells with a jigsaw puzzle piece morphology. One major challenge is to determine what controls the complicated shape change in this cell type and to create computational methods to analyse the geometry of shape change over time.
An earlier study carried out by Professor Szymanski and his colleagues focused on defining the developmental progression of how a pavement cell transforms from a simple polyhedral shape to a lobed morphology. The leading idea at the time was that lobe formation was an iterative process determined by cell size, and as the cell grew and the distance between existing lobes increased, new lobes would form. Through a time-lapse analysis, the Szymanski team found that lobe formation is episodic, and occurs during early cellular development. Once the new lobes have formed, the cell uses this new pattern as a template for future growth that occurs for days. This observation suggests that hormone signalling and other developmental events create a permissive state for polarised growth and these early growth events can have a long-lasting effect on the growth pattern of the cell. The genes and cellular parameters that control this polarised growth process are currently being discovered and analysed.
Another major challenge is to figure out a way to quantify the growth behaviour of cells with such complicated geometric shapes. For a circular or polyhedral cell, it is very simple to graph changes in the size and shape of the cell. However, for pavement cells with an irregular closed geometric shape, how does one measure the formation of a lobe, or measure the growth trajectory of a sub-region of the cell? In a collaboration with the lab of Dr David Umulis – an Associate Professor of Biological Engineering at Purdue – Professor Szymanski’s group created a new tool to quantify the formation of lobes and the local geometry of the cell. The computational tool termed LobeFinder (http://datadryad.org/ resource/doi:10.5061/dryad.cs78t) creates a refined convex hull around extreme points along the cell perimeter, and then measures the distance from the cell perimeter to the convex hull. This method converts the highly irregular shape of a pavement cell into a simple plot of cell shape as a function of location along the cell perimeter. These plots were then used to develop rules to measure lobe formation more consistently than a trained scientist, and much more accurately compared to previous methods. This tool now provides an objective and accurate way to analyse mutant plants to determine if they really do affect the formation of new lobes, or the outgrowth of existing lobes. This is an important first step in generating a reliable genetic pathway that can be used to control cell morphology. The tool also provides a way to graph the timing and location of lobe formation and cell boundary changes. This is important because now proteins that are thought to be involved in the process of lobe formation can be tested directly in microscopy experiments. Do they appear at a cellular location that predicts the location of a new lobe? This tool also allows one to graph how the magnitude and direction of cell growth changes over time. What genes and cellular activities influence these patterns? How do these subcellular and cellular patterns of growth scale to influence the size and shape of a whole leaf? A combined computational and experimental biology approach is creating the knowledge base that is needed for leaf trait engineering.
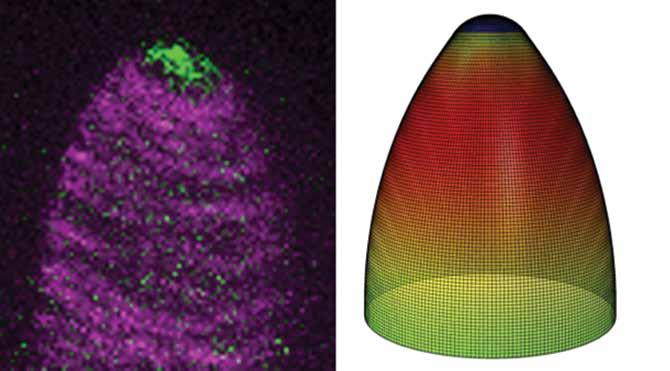
Figure 1. (Left) A live cell image of a real cell with microtubules coloured magenta and the actin filament nucleator ARP2/3 coloured green. (right) A FE computation model of the same cells showing a heat map of the growth patterns of the cell. red (high), blue (low).
Valuable Single Cells: Leaf Hairs and Cotton Fibres
In many cases an important plant trait is determined by a single cell. For example, individual root hair cells generate most of the surface area of a root and affect the efficiency of water and nutrient uptake. On the leaf surface, hair like cells develop from the epidermis, and they can protect the plant against insect attack. Furthermore, the density, shape, and chemical composition of leaf hairs can have a strong effect on how effectively the leaf is protected from herbivory.
Cotton fibre – the most important textile crop in the world – is comprised of individual hair-like cells that develop on the epidermal cells of the seed coat. The value of the fibre is based on its length, diameter and mechanical strength – all of which are the output of the growth properties of a single cell. Unlike pavement cells, these cell types execute a very regular series of cell shape changes as they morph from a hemispherical cell to a highly elongated cell with a tapered tip. Engineering the mechanical properties of a single cell type is much simpler, and it is now possible to understand how individual genes and their encoded proteins can dictate the growth patterns of an isolated cell.
In a study published by Nature Plants entitled ‘Patterning mechanisms of cytoskeletal and cell wall systems during leaf trichome morphogenesis’, Professor Szymanski’s research team discovered new ways in which the internal cytoskeleton of the cells influences shape during growth. In plant cells, the shape of the cell and how it changes are dictated by a tough outer cell wall. The inside of the cell is under very high pressure (about 90 psi, similar to a racing bicycle tire) and the manner in which the cell changes shape is dictated by the local mechanical properties of the wall. The softness of the wall, its thickness and the degree to which cellulose fibres are aligned in the cell wall can all influence how the cell changes shape during growth. The primary purpose of the cytoskeleton is to organise the machinery inside the cell that determine the mechanical properties of the external cell wall. In collaboration with Dr Joe Turner, a materials scientist and engineer at the University of Nebraska-Lincoln, the research team created a finite element computational model of the cell wall of a developing leaf trichome in Arabidopsis (see Fig. 1). The finite element method is a computational technique that treats the cell wall like a pressurised shell, and the shell is divided into discrete sectors with specific mechanical properties. The model is used to identify what mechanical properties of the wall are needed to generate a pattern of shape change that is observed within the living cell. A time-lapsed movie of the growing cell provides a way to validate the accuracy of the predictions from the finite element model (click here for the movie).
The combination of finite element analysis of the cell walls and live cell imaging allowed the team to discover how the microtubule and actin cytoskeletons work together to dictate the shape change patterns of the cell. They analysed the patterning of microtubules and cellulose microfibrils in the cell wall, and found that a specific threshold of fibre alignment transverse to the long axis of the cell must be met in order for the cell to increase in length without any significant radial swelling. The function of actin during plant cell morphogenesis in general is poorly understood, because the actin network is so unstable in most cell types and its patterns are not easily correlated with cell shape. In this study, the team showed that the evolutionarily conserved actin cytoskeleton nucleator – a protein complex termed the actin-related protein (ARP)2/3 complex – generates a stable patch of actin at the tip of the cell (Fig. 1). This actin patch organises the intracellular roadways for material delivery in the cell. Without the patch, the roadways are random, and new cell wall material is not distributed equally in the growing cell wall. When this happens, the cells become swollen and elongation often stops. The team also showed that the region of the cell apex where the actin patch is present is dynamic, and the size of the patch changes as the cell grows to allow it to become more narrow. This process, referred to as ‘tip refinement’ allows the leaf trichome to have a sharp tip that defends against insects. In cotton fibres, tip refinement is a very important trait, as the value of elite cotton varieties is derived, in large part, from the smaller diameter of the fibres. New knowledge from the Arabidopsis leaf hair system is being translated to the cotton fibre system.

Future Research Directions
Professor Szymanski’s work is important because it is generating the knowledge base that is needed to engineer plant cell size and shape. His group has discovered how signalling modules determine the location and timing of actin polymerisation within an individual cell. In the future, these modules can be used in crops such as cotton fibres to engineer fibre quality. The team’s combination of experimental and computational biology is also revealing how the actin and microtubule cytoskeletal systems work together in the cell to control the patterns of growth. These computational methods have the potential to be applied to groups of cells within a tissue, in order to direct strategies to engineer the architecture of plant organs or even whole plants. The next step for their work is to develop integrated molecular and mechanical models of cells in the context of a tissue.
Meet the researcher

Professor Daniel Szymanski
Professor of Agronomy
Department of Agronomy
Purdue University
West Lafayette, Indiana, USA
Professor Daniel Szymanski is a Professor of Botany and Plant Pathology, Agronomy and Biological Sciences at Purdue University, USA. He is currently investigating how plant cells use cytoskeletal systems to pattern the cell wall and the shapes of growing cells. He received his PhD in 1995 from the University of Illinois and since then his research has focused on molecular genetics, live cell imaging, proteomics, and computational modelling of plant cell morphogenesis. Throughout his career, he received multiple awards among which are the Purdue Agricultural Research Award and the Purdue Seeds for Success Award. He is a member of the American Association for the Advancement of Science, the American Society of Cell Biology, and the American Society of Plant Biology.
CONTACT
W: https://ag.purdue.edu/agry/directory/Pages/szymandb.aspx
T: (+1) 765 494 8092
KEY COLLABORATORS
Joe Turner, University of Nebraska-Lincoln
David Umulis, Purdue University
Tae Yoon Kim, Purdue University
FUNDING
NSF (Grant IOS/MCB, Grant No. 1249652 and MCB Grant No. 1121893)
Purdue Agricultural Research Program
National Science Foundation Molecular and Cellular Biosciences (MCB)
National Science Foundation Plant Genome Research Program (PGRP)
REFERENCES
M Yanagisawa, AS Desyatova, SA Belteton, EL Mallery, JA Turner and DB Szymanski, Nature Plants, 2015, 15014.
DB Szymanski, Current Opinion in Plant Biology, 2014, 22, 141–148.
C Zhang, SO Kotchoni, A L Samuels and DB Szymanski, Current Biology, 2010, 20, 1–6.
D Basu, J Le, T Zakharova, EL Mallery and DB Szymanski, PNAS, 2008, 105, 4044–4049.
J-L Qiu, R Jilk, MD Marks and DB Szymanski, The Plant Cell, 2002, 14, 101–118.
DOWNLOAD

{kind=link}